《追寻记忆的痕迹》-完-4.2

作者: [美国] 埃里克·坎德尔
出版社: 中国轻工业出版社
出品方: 万千心理
原作名: In Search of Memory: The Emergence of a New Science of Mind
译者: 罗跃嘉
出版年: 2007-1
页数: 323
定价: 35.00
装帧: 平装
丛书: 心理学导读系列
ISBN: 9787501956500
简介:
力图从细胞和分子的生物学角度来研究认知心理加工。认知神经科学。
作者:
埃里克·坎德尔(Eric R. Kandel),犹太裔神经科学家。1929年出生于奥地利维也纳。1952年本科毕业于哈佛大学历史与文学专业,1956年在纽约大学获得医学博士学位,目前担任美国哥伦比亚大学教授和霍华德·休斯医学研究所资深研究员。他一生致力于神经科学的前沿研究,因在记忆存储的神经机制研究中做出重大贡献,于2000年获得诺贝尔生理学或医学奖。其自传《追寻记忆的痕迹》,曾荣获2006年度《洛杉矶时报》图书奖和2007年度美国国家学院知识传播奖;根据自传改编而成的同名纪录片,曾荣获2010年德国巴伐利亚电影奖最佳纪录片奖。
译者序中,知道这个版本是大学实验室导师,率一众学生翻译的作品,也强调了集体校对的环节。但对这种翻译的形式,也是发自内心的不认同。或许只有这种方法,才能让这类科普书籍尽快发行吧。
这本书上,又有之前借阅者的铅笔划痕,还有一张塑料片麻将的书签。很厌烦,又得拿橡皮擦拭。
这两个多月,每批阅读的书籍中,都有跟犹太有关的内容。有些是内容的关联性的接续阅读,但这本不是,未曾想到作者是犹太裔。
作者用几章的笔墨在描写儿时维也纳,描绘犹太民族。这个民族的人真是适应力顽强,且有高阶的目标和憧憬——更多的教育和文化。
在第2章,作者对维也纳反犹的描述说,文化思想,道德观念不是判断人性的标尺,人们对异族的排他,嫉妒和复仇才是最丑恶的。无独有偶,《希腊罗马名人传》的传记中,几乎都有描写因为嫉妒对主人公的迫害。嫉妒,才是不安分,不安定的始作俑者。很少有人愿意正面这种情绪的产生和恶果。
译者们,真是太不讲究了,相关人物名字都用英文,怎么不整本书都英文呢,别翻了。人物关联的其他书,没办法辨别。这些国外的科学家真是,翻译觉得该有中文译名就有,没有就直接字母。怎么能这么随意呢。
1-3翻译很流畅。4翻译一般。5内容讲得很明白。6这一章,堪比左右互搏,信息量比较大,累脑子。
记忆存在哪,怎么存的,真是个问题。
第9章,看到作者对实验室合作伙伴Alden的描述,发现他仅活到46岁,感到很惋惜,这可能是科研工作者最好对年华。世界进程是这样的人改变的,可却没给他更多对时间。
这本书,我理解甚至都不算是一本科普,因为下面的笔记,没准那条就是不对的,跟后面的内容,或者现在的内容矛盾,作者用自传的方式,在写一部跟自己科研领域相关,尤其跟自己所在年代相关的传记研究史。
以下笔记信息不像地球是圆的那么肯定。我当知识的摘抄记录,甚至下一段就被告知是不对的。哎,知道科研工作者的遭遇经历和发展进程,但前提不给。特别不喜。都想给这书3分了,你会发现用心记的东西,是发展中被淘汰的,后面提及的也不见得是对的。到处都是这种“真真假假”的信息穿插,阅读体验非常糟,完全不是科普读物的逻辑布局,所以说是传记,传记就没必要按照知识来进行笔记了。
书中提及了还原法,简单说就是微观入手的方法论,特意在网上搜了科普信息:https://blog.sciencenet.cn/blog-279293-1154766.html 讲得很明白细致,还有整体论和新整体论。
达尔文曾经提到的他的“对于自然科学的热爱……实际上很大程度上出于想要得到同行尊重的野心”。
第一卷
心理分析是一门通过分析个人早期记忆和经验来理解人们的动机、思维和行为的非理性根源的学科。
有一个是我困扰的结论是,一个社会的文化素质不是其是否尊重人性的可靠标准。文化本身并不能改变人们的偏见和思维。消灭不属于自己团体的人的欲望或许是一种本能反应,因此可能在任何凝聚性强的团体内出现。
——犹太人的生存能力和赚钱的能耐真是了不得。
国家社会主义(纳粹主义)。
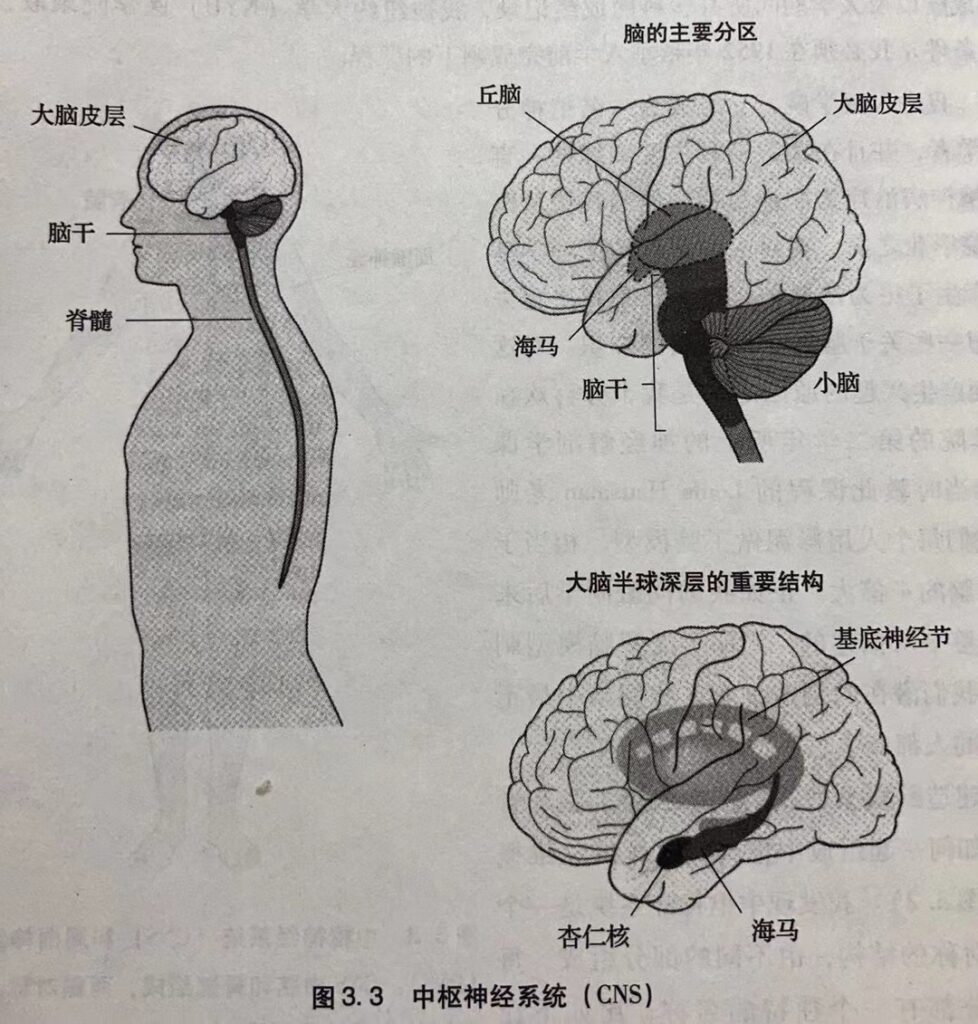
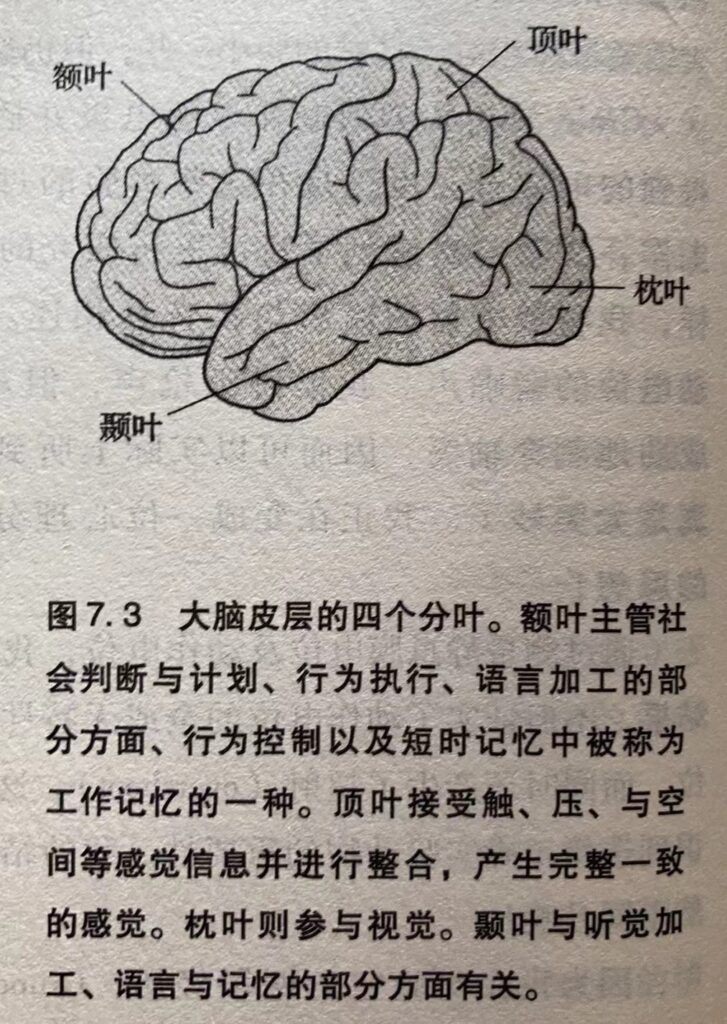
脊髓向上与延伸至脑形成脑干(图3.3),它负责将感觉信息传递到更高级的脑区,然后又将运动指令从这些区域传送到脊髓。 脑干还调节和控制着注意。在脑干上面依次是下丘脑、丘脑和大脑两半球。大脑半球的表面覆盖着一层高度褶皱的表层–大脑皮层。大脑皮层执行知觉、动作、语言和计划等高级大脑功能。大脑的深层结构由三部分组成:基底神经节、海马和杏仁核(图 3.3)。基底神经节负责协调运动,海马参与记忆储存,而杏仁核则负责协调某种情绪状态下的自发和内分泌反应。
Denise却坚持钱财并不是问题。她的父亲在我遇到她的一年前去世,生前曾建议她与一个清贫的知识分子结婚,因为这种人会将学识视为高于一切,会去努力追求和实现崇高的学术目标。
第二卷
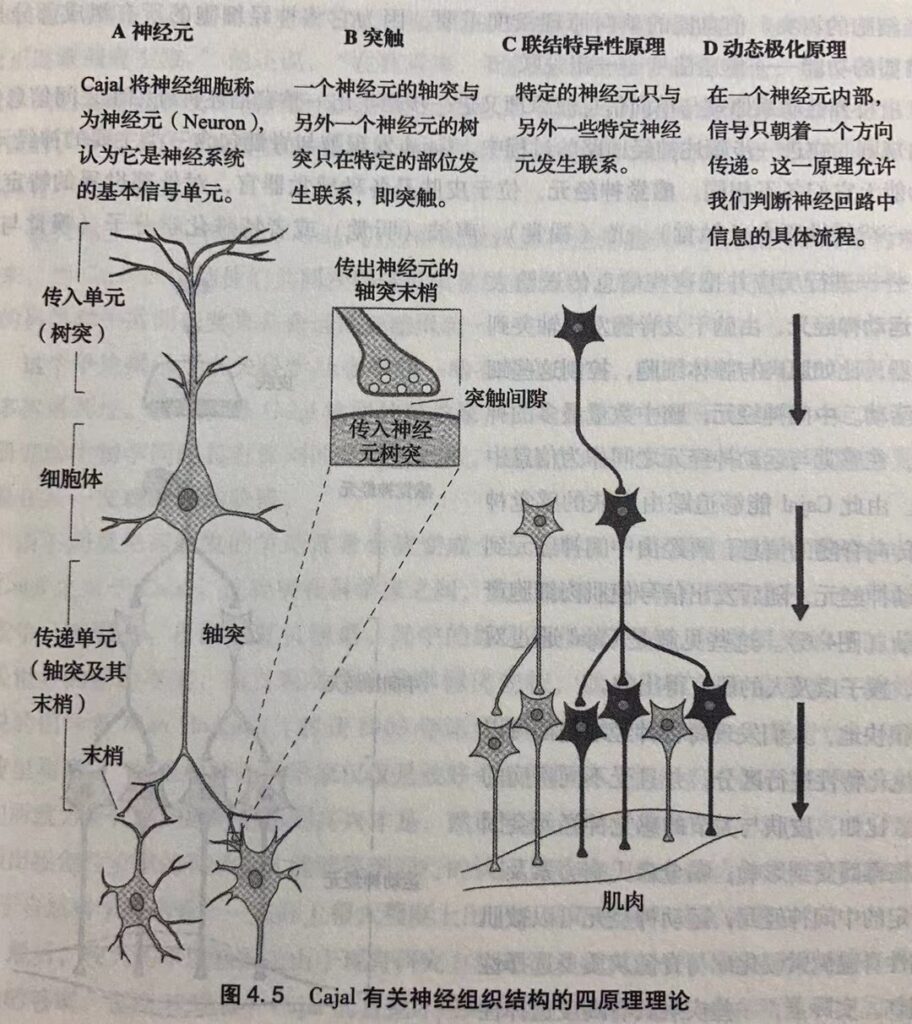
神经细胞生物学以三个原理(神经元理论、离子假说、突触传递化学理论)为基础,始于20世纪的上半叶,而今成为理解脑功能的核心。
神经元理论(细胞理论应用于脑):阐述了神经细胞(或者神经元)是脑的基本信号单位的根本组成部分。——西班牙解剖学家Cajal(1852-1934)提出。
与人体其他结构简单的细胞不同,神经细胞具有极端不规则的形状,并且被众多的极其精细的延伸部分(当时称为突触)所包围。
1890年Cajal提出神经元理论四个原理:
原理一:神经元是脑的根本机构与功能单位,即基本的组成部分与基本的信号传递单位。
原理二:神经元树突之间的联系只发生特定部位(突触),两个神经元之间的突触有一个小间隙相隔。
原理三:每个神经细胞只与特定的神经细胞形成突触而产生联系。
原理四:动态极化,神经回路中的信号只向一个方向传递。
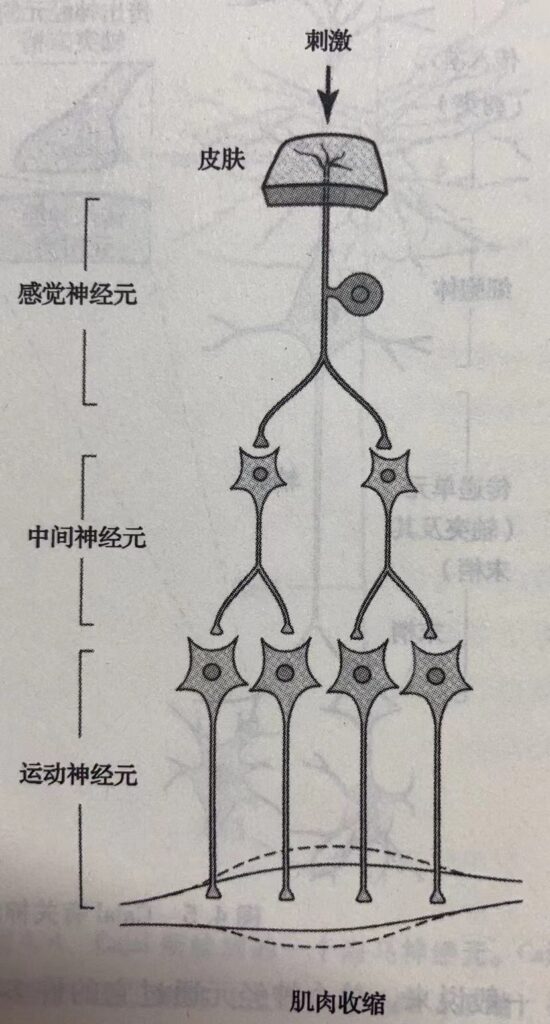
由特异性联系原理与单向信息流原理又进一步衍生出一整套描述神经细胞之间信息传递的规则。在进一步阐述神经回路的过程中,Cajal 发现脑与脊髓包含三类主要的神经元,在功能上它们各不相同。感觉神经元,位于于皮肤及各种感觉器官,对外部世界的特定刺激——机械性压力(触觉)、光(视觉)、 声波(听觉)或者特殊化学分子(嗅觉与味觉) ——进行反应并把这些信息传送给大脑。运动神经元,由脑干及脊髓发出轴突到效应器,比如肌肉与腺体细胞,控制这些细胞的活动。中间神经元,脑中数量最多的神经元,在感觉与运动神经元之间作为信息中转站。由此Cajal 能够追踪由皮肤的感觉神经元传向脊髓的信息,再经由中间神经元到达运动神经元,随后发出信号使肌肉细胞产生运动。
离子假说:则集中于神经细胞内部的信息传递。它描述了单个神经细胞产生电信号(即动作电位),并在特定细胞内沿着相当长的距离进行传导的机制。
神经系统以一种低速(约30米/秒)的主动传导来保证从大脚趾产生的信号可以大小不变地到达脊髓。
强度产生于动作电位的激发频率。感觉的持续时间是由产生动作电位的时间长短决定的。碰撞手肘,可激发每秒上百个动作电位。不同刺激,并非依赖于动作电位的不同。所有的(神经)冲动都差不多。每一种感觉都有特定的神经通道传导,特定的信息专门由传导该信息的神经回路中的神经元传导。
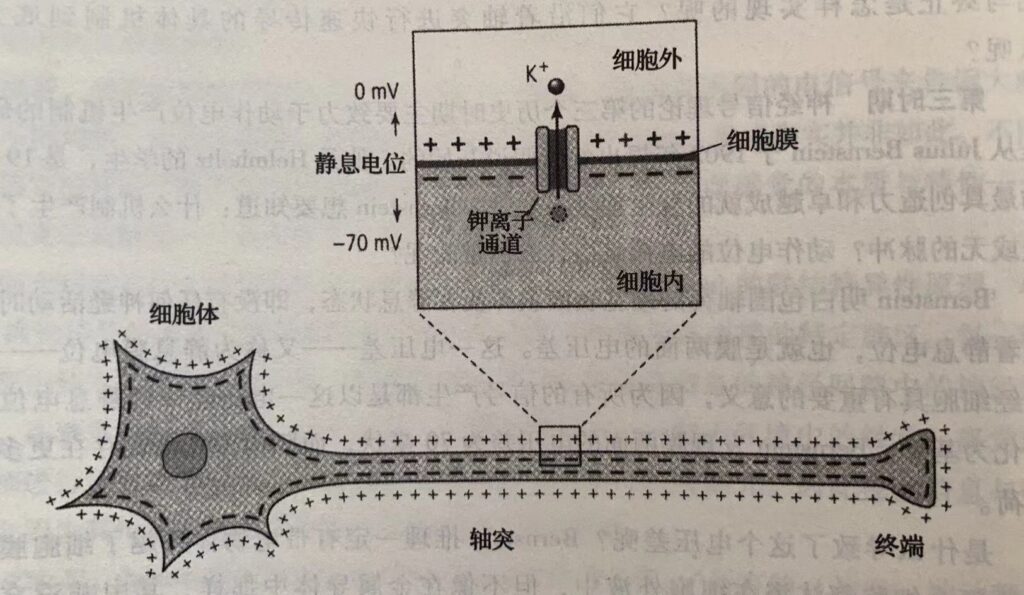
Bernstein 明白包围轴突的细胞表面膜即使在静息状态,即没有任何神经活动时也存在着静息电位,也就是膜两面的电压差。这一电压差–又称为静息膜电位–对于神经细胞具有重要的意义,因为所有的信号产产生都是以这一电压差(即静息电位)的变化为基础。Bernstein发现膜两面的电压差为70毫伏,而膜内侧比外侧存在更多的负电荷。
静息电位为-70毫伏,由钾离子通过离子通道的移动而产生的。动作电位的波幅为110毫伏,动作电位升支,其上升及最大波幅取决于细胞外液中钠离子浓度。动作电位降支,则与钾离子浓度有关。当细胞膜受到充分刺激后,钠离子以每秒千分之一的速度移向细胞内,从而将内部的电压由-70毫伏提升到40毫伏进而产生动作电位上升段。而增强的钠内流立即引起钾外流的增强,从而产生动作电位的降支,细胞内部电压又回到初始状态。钠、钾离子的电压门控通道实际上是一些蛋白质。
20世纪中叶被证实是错误的。
突触传递的化学理论:则主要论述了神经细胞之间的信息传递。一个神经细胞通过释放化学信号(即神经递质)与另外一个神经细胞联络;第二个神经细胞识别信号并通过位于细胞膜表面的、被称为受体的特殊分子进行反应。这三个原理都聚焦于单个的神经细胞。
自主神经系统的神经细胞体聚集成簇,称为周围自主神经节,位于脊髓与脑干的外部,是周围神经系统的一部分。自主神经系统控制重要的非随意运动,比如呼吸、心率、血压及消化。
现称那种化学物质为神经递质,由突触间隙向靶神经元移动,最后被靶神经元表面膜上特殊受体识别并捕获。
迷走神经减低心率;交感神经提高心率。
迷走神经释放的物质实际上是一种简单的化学物质乙酰胆碱,与特殊受体结合是心率降低。
神经通路上的细胞无时无刻不在接受众多的突触信号,有抑制性的也有兴奋性的,但结果只有两种情况:产生动作电位或是不产生动作电位。一个神经细胞最为基本的任务即是整合:整合它接收到的来自突触前神经元的兴奋性或抑制性突触电位,仅仅当兴奋性信号的总和超出抑制性信号的总和至少达到一个最小阈值时才产生动作电位。
中枢神经系统的突触电位是化学启动(化学传递)还是电启动(电传递)的?
乙酰胆碱由运动神经元释放的直接证据,并因此可适合于突触电位的所有阶段。乙酰胆碱快速地弥散到突触间隙中,然后与位于肌细胞上的受体迅速结合。后来,人们发现乙酰胆碱受体原来是一个有两个主要成分的蛋白质:一个乙酰胆碱结合成分和一条离子通道。当乙酰胆碱被识别并与受体结吉合时,离子通道随即开放。
Katz继续证实这条新发现的化学递质门控的离子通道与电压门控的钠与钾通道有两点区别:它们只对于特殊的化学递质反应, 既允许钠也允许钾通过。钾与钠离子通道的同时开放将肌细胞膜的静息电位由-70 毫伏提高到接近0毫伏。就如 Dale 曾经预言的那样,尽管突触电位由化学物质引发,但也非常迅速。当突触电位足够大时,便产生动作电位从而使肌细胞收缩(图6.2)。
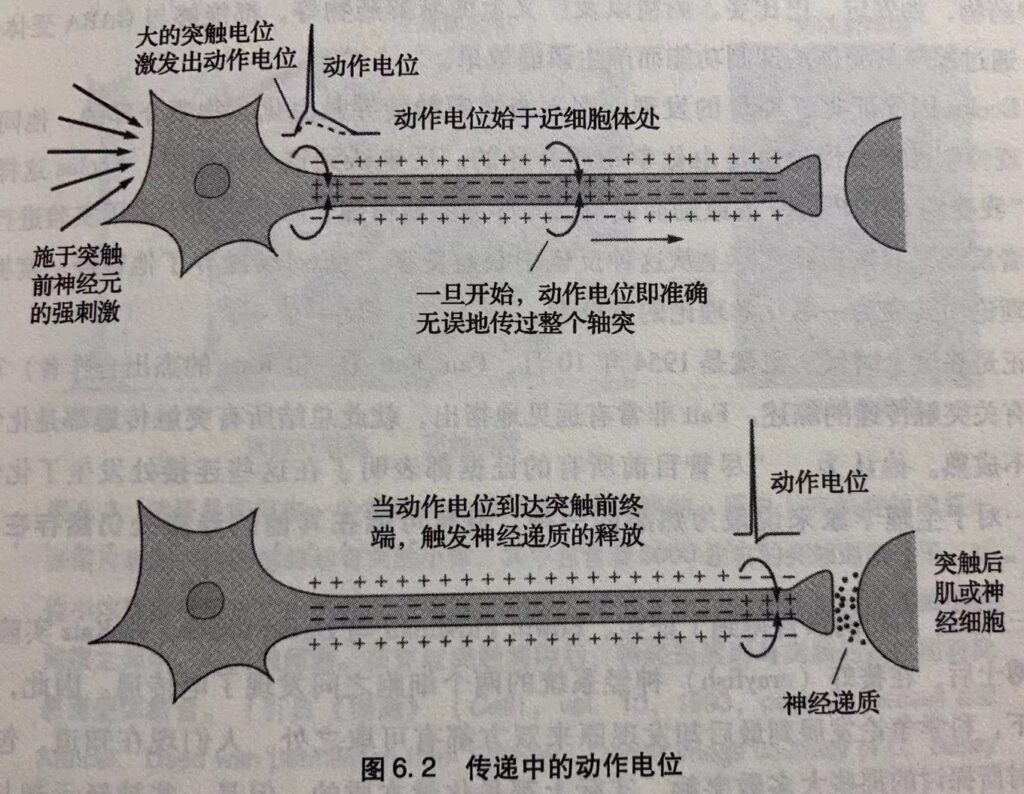
运动神经元同时接受由不同神经递质作用于不同受体而产生的兴奋性与抑制性信号。运动神经元中突触前神经元释放兴奋性神经递质将膜静息电位由-70毫伏提高到-55 毫伏,即引发动作电位的阈值;而抑制性神经递质则将膜静息电位由-70毫伏减低到-75毫伏,因而使动作电位更难激发。
现在人们知道脑内最主要的兴奋性神经递质是谷氨酸(amino acid glutamate),而最主要的抑制性神经递质则为γ-氨基丁酸(GABA,gamma-aminobutyricacid)。许多镇静药物,如安定、巴比妥、酒精以及广义上的麻醉药物等,都能够与 GABA 受体结合,通过提高其受体的抑制功能而产生镇静效果。
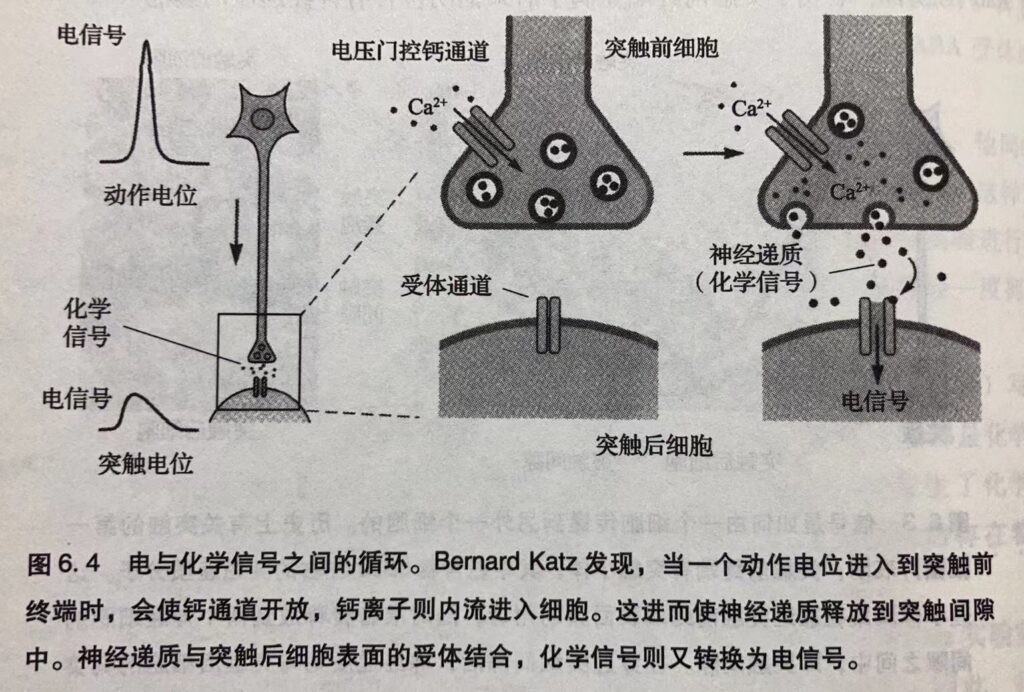
在这一不同寻常研究的最后阶段, Katz将他的注意力由靶细胞的突触电位转向了信号细胞的神经递质释放。他想要知道突触前终端的电事件,即动作电位是如何引发化学递质的释放的。在这一研究领域, 他主要做出了两个重大发现。首先,当动作电位沿着轴突传到突触前终端时,电压门控的离子通道开放,钙离子流入。突触前终端钙离子的流入进而导致了神经递质的释放。 因此,对于发出信号的细胞来说,由动作电位打开的电压门控钙离子通道开启了将电信号转换为化学信号的程序。同样,对于接收信号的细胞来说,递质门控的离子通道则将化学信号再转换为电信号。
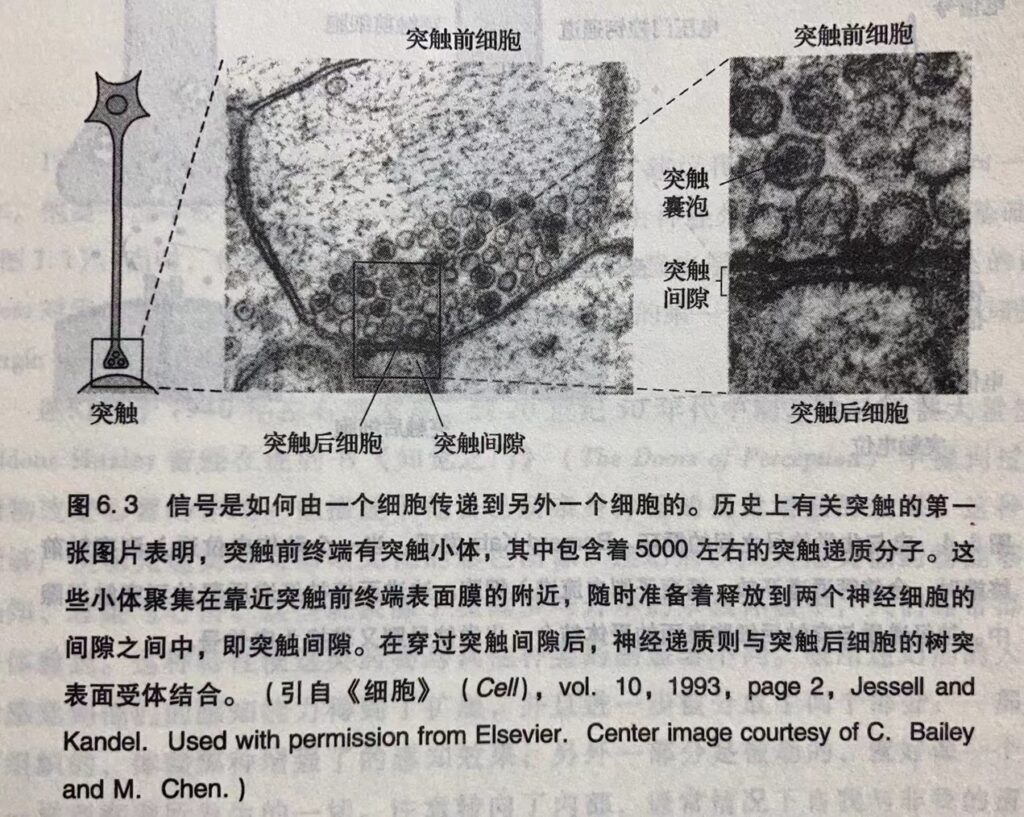
其次,Katz发现轴突末端释放乙酰胆碱之类神经递质时,并不是以单个分子的形式而是以小的互相独立的小囊泡(每个包含大约5000个分子)的形式。Katz将这些小囊泡称为量子(quanta),并且推理它们每个都是由膜包裹的小囊,后来他称为突触囊泡(synapticvesicle)。1955年,Sanford Pala y与George Palade由电镜下获得的图像证实了 Katz 的预言,表明了突触前终端充满了后来证明包含有神经递质的囊泡(图6.3)。
迷幻剂以及其他类似药物能够改变感知、思维与心情。这些现象除了在睡李梦与升华的宗教体验中,人们通常都不能够体验到。这种特性使这类药物与其他和种类药物显著不同。服用迷幻剂的人通常会感觉到他们的感知能力得到了扩展,主并且进一步被分成了两个部分:一部分是有组织的,体验那种增强了的感知效果; 另外一部分是被动的,就好像一个旁观者一样观察着所发生的一切。注意转向了了内部,通常情况下自我与非我的清晰分界不再存在,这使服用迷幻剂者产生一和中神秘的感觉,好像自己是宇宙中的一部分。大多数服用者的认知扭曲通常表现为为幻视;而另外一些服用者则产生一种类似精神分裂样的反应。
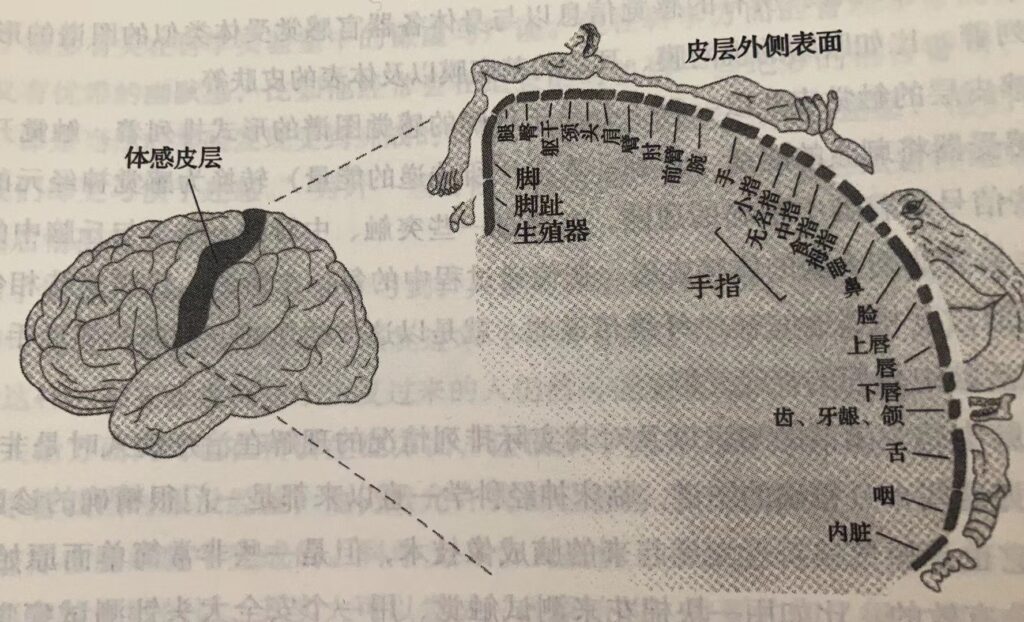
一种特殊类型的癫痫发作当这种癫痫发作时,麻木及烧灼或刺痛感首先由身体一处开始然后遍及全身。比如,麻木可能首先由指尖开始,过几分钟后则延伸到手部,然后是胳膊、再绕过肩膀到达后背,最后延及同侧腿。这种感觉异常出现的序列就可以用脑中体感区的感觉图谱来解释:由脑内的阵发性异常电活动而引发的癫痫发作,由侧面体感区表征手的区域开始,然后延及皮质中线表征腿的区域。
大脑皮层有两个重要特征。第一,尽管它的两个半球看起来彼此相似,但实际上它们在结构与功能上都有显著的不同。第二, 每个半球分别负责感觉与移动对侧肢体。因此,由左侧肢体(比如说左手)发出到达左侧脊髓的感觉信息最终投射到右侧大脑皮层。同样,右半球运动区其实负责左侧肢亿本的运动。
“我们用左脑讲话!”
这种由专门化区域所构成的大而互相联系的神经网络(比如负责语言的区域)正是人们具有种种天衣无缝心理功能的首要前提。
脑回路的内在冗余性。许多感觉、运动以及认知功能都由不止一条的神经回路来实现,同样的信息同时由不同脑区平行加工。当一个区域或者通路受损,其他的可以至少部分地代偿受损的功能。当代偿发生时,行为缺陷则不明显,于是研究者就很难将一种行为与某一处脑损伤相联系。
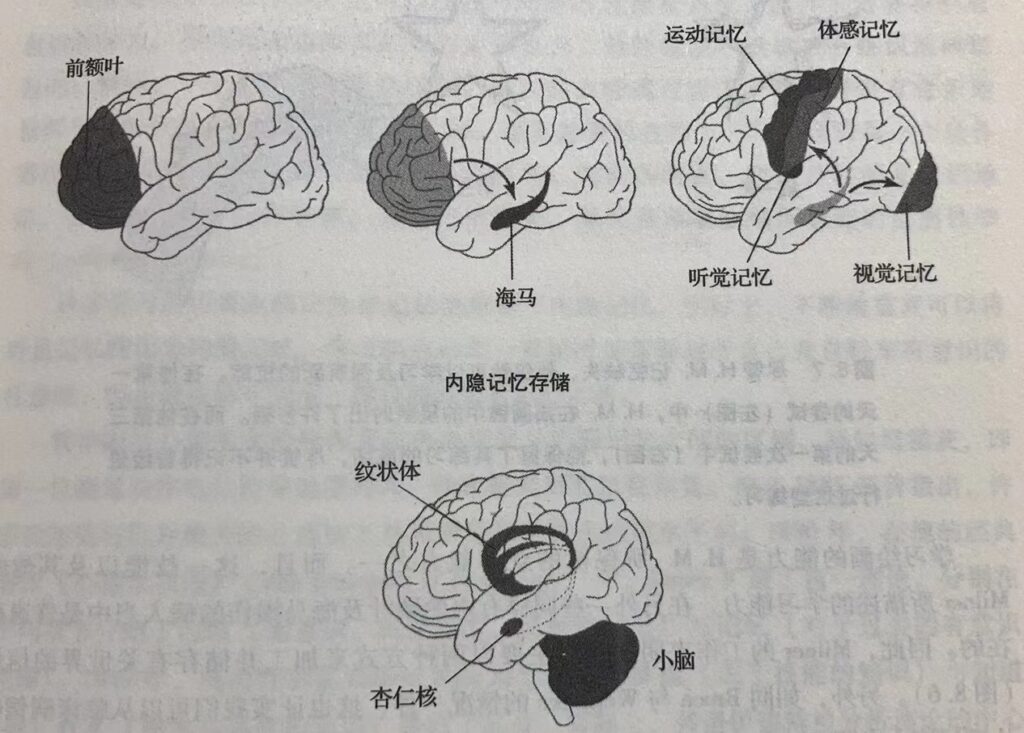
通常所认为的有意识记忆,现在实际上被称为外显记忆,或者是陈述性记忆(declarative memory)。这是由Squire及Schacter首先提出的。它是对于人物、地点、物体、事实以及事件等的有意识回忆,这些都是H.M.所欠缺的记忆。无意识记忆现在被称为内隐记忆(implicit memory),或者是程序性记忆(procedural mem- ory)。它构成习惯、敏感化、经典条件反射、感知与运动技能,比如骑自行车或者打网球等的基础。这即是H.M.尚且保存的那种记忆。
内隐记忆并非是一个单一的记忆系统,它是一组包括大脑皮层下面几个不同脑结构的加工集合(图8.6)。例如,与事件相关的感觉联系(害怕或者高兴) 就主要与一个叫做杏仁核的结构有关。形成新的运动或者是认知习惯,与纹状体有关,而学习新的运动技能或者是协调活动则与小脑有关。在包括无脊椎动物的最简单动物中,与习惯、敏感化以及经典条件反射有关的内隐记忆甚至可以储存在反射通路中。
内隐记忆具有自主性。它可以在执行动作时直接被回忆,而并不需要我们有意识的努力。尽管经验会改变认知与运动能力,但经验是不能够被有意识地回忆起的。例如,一旦你学会了骑自行车,你只要去骑就可以了。你不需要有意识地指挥自己的身体:“现在伸出我的左脚,现在是我的右脚……”如果对每一个动作都过多地关注,我们反而可能会摔下自行车。说话的时候,我们自动而无意识地说。这就是一类由行为学家,诸如巴甫洛夫、桑代克和斯金纳所研究的反射性学习(reflexive learning)。
许多学习经历既依赖于外显记忆也依赖于内隐记忆。实际上,不断地重复可以将外显记忆转化为内隐记忆。学习骑自行车一开始时是需要对于自己及自行车有意识的注意的,但是最后骑车变成一种自动而无意识的运动。
记忆的细胞学研究必须回答几个主要问题:当我们学习的时候大脑内部发生了怎样的变化?不同种类的学习是否会产生不同种类的变化?记忆存储的生化机制是怎样的?这些问题都在我的脑中盘旋着,但这样的问题很难轻易地被转化成有用的实验。
“Jim从来不将深入思索与刻苦工作混淆。”
第三卷
三种类型内隐学习:习惯化、敏感化、经典条件反射。
习惯化是最简单的学习形式,通过习惯化,动物学会识别无害的刺激,这样就能在确保自己安全的情况下对它们置之不理。使人们能够在相对嘈杂的环境中工作。使我们对钟表的声音以及心跳、胃蠕动等躯体感觉习以为常。习惯化是学习识别反复发生的无害刺激的过程。习惯化另一个作用,消除了不必要的或过激的防御反应。
习惯化不只局限于逃跑反应:性活动也有习惯贯化现象。当雄鼠遇到一只雌鼠后,在最初的一到两个小时内,它们可以交配六到七次。 但是在以后的30分钟或更长时间内,雄鼠不再对性活动有兴趣,交配频率明显下降。请注意,这种现象是习惯化,而不是疲劳造成的。因为如果这只雄鼠面临一只新的此堆鼠,交配活动将再度活跃。
——老鼠都会腻,何况人。老鼠都好新鲜,何况人。
习惯化还可以作为一种测量工具,我们可以借助它判断个体对物体的熟悉程度。
习惯化是教动物学会忽略某刺激,与之相反,敏感化是一种恐惧习得形式:威胁刺激发生后,动物立即变得敏感,对几乎一切刺激都投入更多的注意,并产生更强的反应。例如,向动物足部施加电击后的短时间内,动物对铃声、音调或轻柔的触摸等刺激都表现出过激的退缩和逃跑反应。
最后,我试图模拟负性经典条件反射 这种经典条件反射教动物将不愉快的刺激(如电击)和不诱发反应的普通刺激联系起来。中性刺激必须总是在负性刺激之前呈现,前者似乎预示着后者的发生。例如,巴甫洛夫用施加在狗爪上的电击作为负性刺激。电击诱发出动物的恐惧反应:狗抬起腿并向后缩。巴甫洛夫发现,将电击和铃声配对呈现多次后总是铃声在前,电击在后-狗听到铃声就会缩腿,即使在没有电击的情况下也是如此。因此,负性经典条件反射是一种关联性恐惧习得形式。
负性经典条件反射和敏感化的类似之处在于,一个感觉通路的活动增强了另一个感觉通路的活动。然而,二者是不同的,它们的不同点表现在以下两个方面。首先,经典条件反射使一对相继发生的刺激形成联系。其次,经典条件反射只增强动物对某一个中性刺激的反应,而敏感化使动物对整个环境的反应增强。
——一群最聪明的大脑聚在一起,科学家聚团可真是令人赞叹的环境,科研氛围。
首先,突触联系强度的变化足以改变原有的神经网络结构及其信息加工能力。
其次,不同形式的学习可以以完全相反的方式改变两神经元间的突触联系——强化或弱化。具体来说,习惯化学习弱化了突触联系,而敏感化学习和经典条件反射强化了突触联系。突触联系强度的改变是学习和短时记忆的细胞机制。
第三,短时记忆存储的时间长短取决于突触强化或弱化的时间长短。
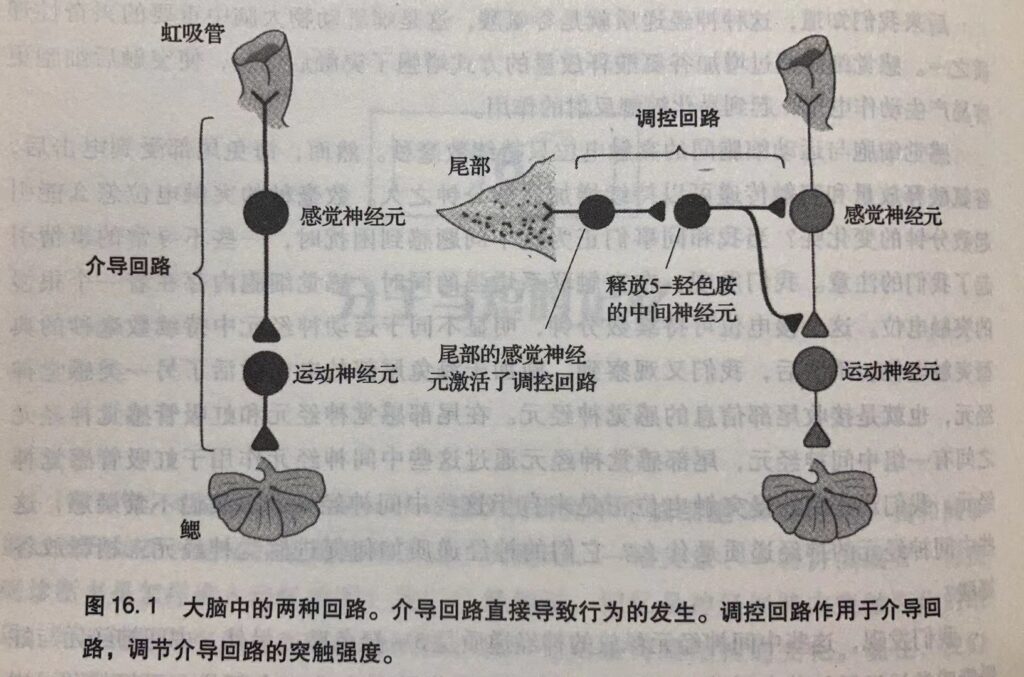
第四,突触强度的调控方式分为同突触和异突触两种。究竟采取哪种方式,取决于学习激活了哪个神经回路——是介导回路,还是调制回路。
艾宾浩斯遗忘曲线。遗忘分为两个阶段:学习后的一个小时里,记忆量急速下降,形成陡峭的遗忘曲线;第二阶段,记忆缓慢速度丢失,持续时间为一个月左右。
临场证据都说明,记忆储存的早期阶段是不稳定的,容易受到干扰。
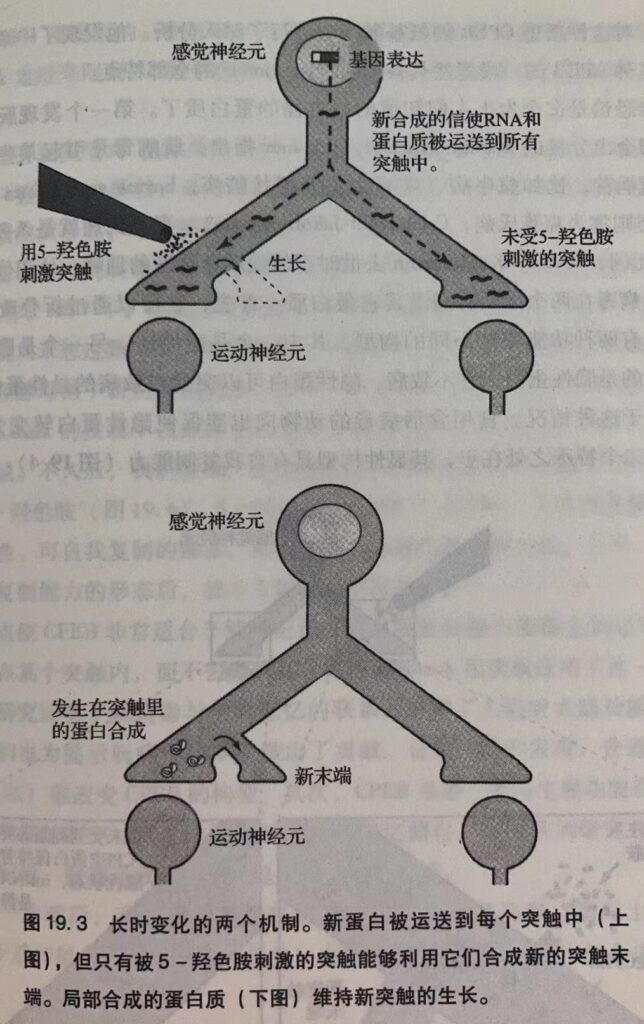
记忆包括一个持续数分钟的短时记忆和一个持续数天以上的长时记忆,前者通过固化过程转化为后者,固化过程需要合成新的蛋白质。
大脑中突触的数目不是一成不变的,学习能够改变突触的数目。敏感化学习使神经元长出新的突触末端,习惯化则使神经元现有的突触末端数目减少。学习能够激活无活性的突触,也能让有活性的突触失活。
短时记忆是突触功能变化的结果,长时记忆还需要结构的变化。
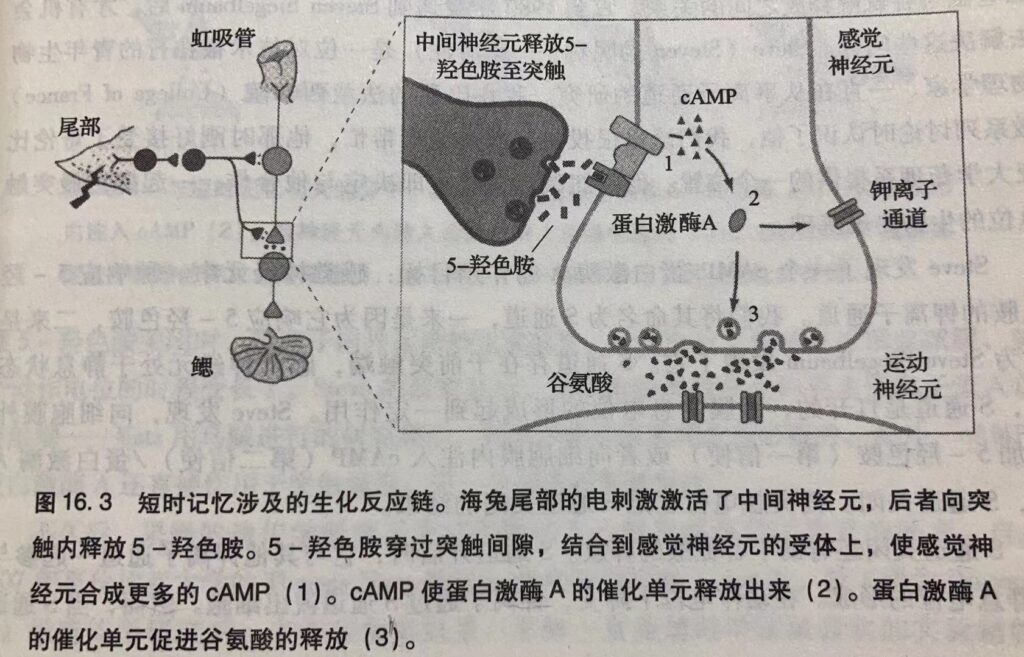
神经递质就是谷氨酸,感觉细胞通过增加谷氨酸释放量增强了突触后电位,使突触后细胞更容易产生动作电位。
上述发现让我们认识到,学习和记忆中存在着两种重要神经回路 :介导回路 和调控回路 。介导回路直接产生行为,其本质是康德式的,其神经结构是由遗传和发育因素决定的。在我们的研究中,介导回路由虹吸管的感觉神经元、中间神经元以及鳃运动神经元组成。介导回路在学习过程中扮演学生,需要学习新的知识。调控回路的本质是洛克式的,它在学习过程中扮演老师。调控回路不直接参与行为的产生,但学习时,它会通过感觉与运动神经元之间的突触对行为进行微调(这是一种异突触调控)。电击海兔尾部时,调控回路指导海兔注意虹吸管上的刺激,因为虹吸管上的刺激可能威胁到动物的生命安全。看起来,调控回路是造成海兔过激反应的原因。我们将在后面的内容中看到类似的情况–在较复杂的动物中,调控回路是记忆的基本神经成分。
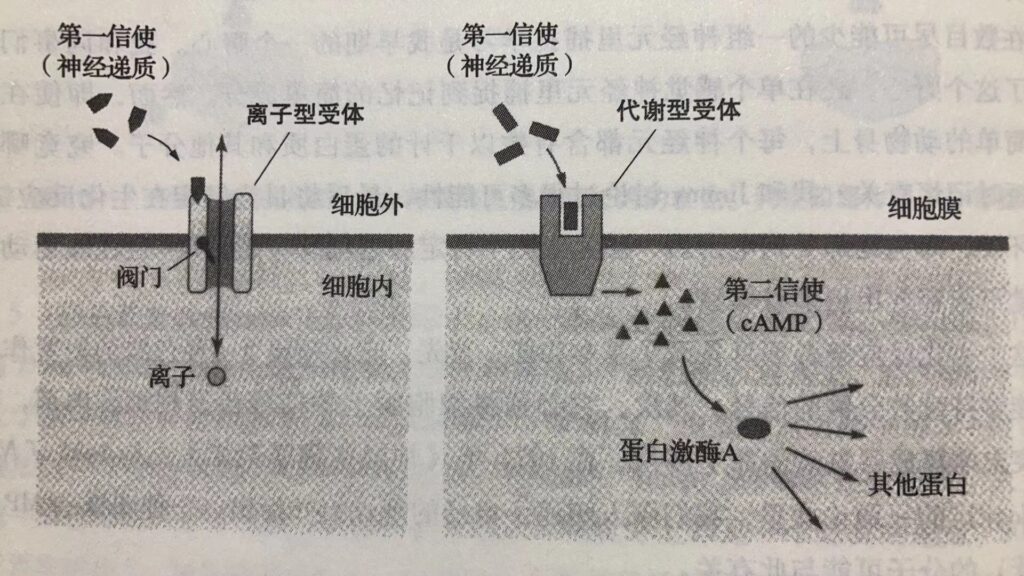
长时记忆需要合成新的蛋白质,细胞必须合成新的结构蛋白。而短时记忆不需要。
Cajal 指出,大脑中的神经元彼此相连,构成许多特定的神经通道。这种特异性在海兔身上得到了证实:特定的神经元以特定的方式彼此相连,构成了缩鳃反射的神经通道。Jimmy告诉我,这种特异性也存在于分子水平也就是说,细胞内的各种分子以特定的方式彼此结合,构成细胞的基本功能单位。生化学家发现,细胞内的各种分子按照一定的次序发生反应,并以这种方式传递信号。信息沿着这条传导途径从细胞表面传递至细胞内部,正如信息沿神经纤维从一个神经元到另一个神经元一样。另外,这条传导途径是“无线”的,由分子介导。在细胞中飘浮的分子识别并结合到特定的目标分子上,改变后者的活性。
即使在海兔这么简单的动物身上,每个神经元都含有数以千计的蛋白质和其他分子。
——嘿嘿,翻译这本书的人,不知道薛定谔的中文译名。甚至不对什么人写了这本书感兴趣,或者不知道如此大名鼎鼎,或者死活都得名字原文。
1953年4月25日,JamesWatson和FrancisCrick在《自然》上公布了 DNA的结构模型。在生物学家Rosalind Franklin和Maurice Wilkins拍摄的X-射线衍射图的帮助下,Watson和Crick推断出,DNA由两条长链构成,两条链相互缠绕,形成螺旋型的结构。他们知道,DNA的双螺旋结构是由4种反复出现的核苷酸构成,它们含有不同的碱基,分别是腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)和胞嘧啶(C)。 Watson 和 Crick 设想,基因中编码遗传信息的元素可能就是这4种核苷酸。后来他们发现,DNA的双链是互补的,一种碱基只能特异性地与互补链上的另一种碱基结合,形成一个碱基对。具体地讲,腺嘌呤只能与互补链上的胸腺嘧啶结合,而鸟嘌呤只能与互补链上的胞嘧啶结合形成碱基对。一串碱基对牢牢地将两条DNA链联系在一起。
Watson 和 Crick 还进一步阐述了蛋白质合成的机制。他们推断,DNA的碱基序列中携带着蛋白质的密码,指导着蛋白质的合成 复制时,基因携带的密码通过合成互补链的方式被“读出”。然而,正如后来的研究显示的那样,在蛋白质合成过程中传递遗传密码的分子并不是DNA,而是一种叫做信使RNA(核糖核苷酸)的分子。像DNA一样,信使 RNA也是由4种核苷酸组成的。其中三种核苷酸的碱基与DNA 完全一样–腺嘌呤、鸟嘌呤和胞嘧啶,但胸腺嘧啶被尿嘧啶取代。DNA的两股链彼此分开后,只有其中一股被转录成为信使RNA。细胞再根据信使RNA的碱基序列合成蛋白质。就这样,Watson 和 Crick 阐明了分子生物学的中心法则:DNA制造 RNA,RNA制造蛋白质。
20 世纪70 年代末,哈佛大学的Walter Gilbo ert 和剑桥大学的Frederick Sanger发明了一种快速检测DNA 碱基序列的新技术。这是一项很了不起的发明。从此,人们就能相对容易地读出某段 DNA的碱基序列,并推测出这段基因编码的蛋白质。借助这项新技术,科学家们观察到,相同的DNA序列出现在不同的基因里,为各种不同的蛋白质编码完全一样或相似的区域。人们将这些可识别I的区域称为结构域。相同的结构域行使相同的功能,无论它们出现在什么样的蛋白质中。现在,科学家们已经能够根据基因的碱基序列大致推测出蛋白质的某些重要信息 比如,这个蛋白质是不是激酶、离子通道或者受体。此外,通过比较蛋白质的氨基酸序列,还能推断出两种蛋白质的相似程度。
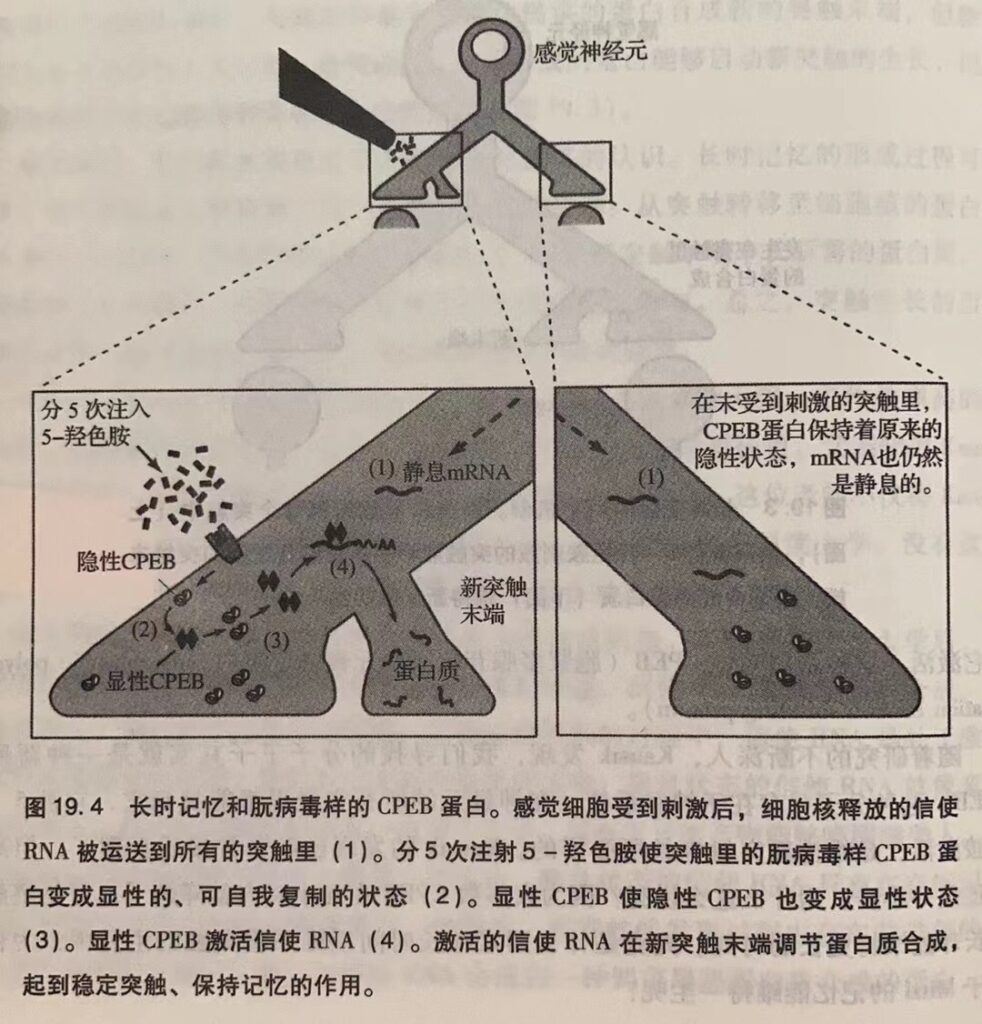
显然,具有自我复制能力的分子可以无限期地存在于突触里调节局部的蛋白质合成,维持新的突触生长。
过去我曾设想过朊病毒与长时记忆的关系,但都不是什么认真的想法。我很了解 Prusiner的工作。1997年,他凭借对朊病毒和朊病毒病的研究获得了诺贝尔生理学或医学奖。尽管我从未想过新型CPEB可能是一种朊病毒,但Kausik的话让我备感振奋。
朊病毒在酵母菌中研究得比较多。然而,在Kausik之前,没人知道这种蛋白质的正常功能。Kausik 的发现不仅加深了人们对学习和记忆的认识,还开辟了一块新的生物学研究领域。不久后,我们发现,在缩鳃反射的感觉神经元中控制CPEB构型变化的物质正是5-羟色胺(图19.4)。5-羟色胺使CPEB从无活性、无自我复制能力的形态转变为有活性、可自我复制的形态,从而维持局部蛋白质合成过程。另外,一旦CPEB变成有自我复制能力的形态后,就不太容易再变回去了。
上述特点使CPEB非常适合于储存记忆,其自我复制能力使信息得以有针对性地、持久地保留在某个突触内,而不去影响其它突触。Kausik很快就证明了这一点。
我们的研究证明了朊病毒与长时记忆的联系,扩宽了人们对大脑功能的认识。不仅如此,我们也为揭示朊病毒的本质做出了贡献。首先,我们发现,普通的生理信号(如5-羟色胺)能改变CPEB的构型。其次,CPEB是第一个与生理功能有关的朊病毒 –具体到我们的研究,就是维持突触易化和记忆储存。在以往的研究中,有自我复制能力的朊病毒一般都是致病因子或是无活性的蛋白质。
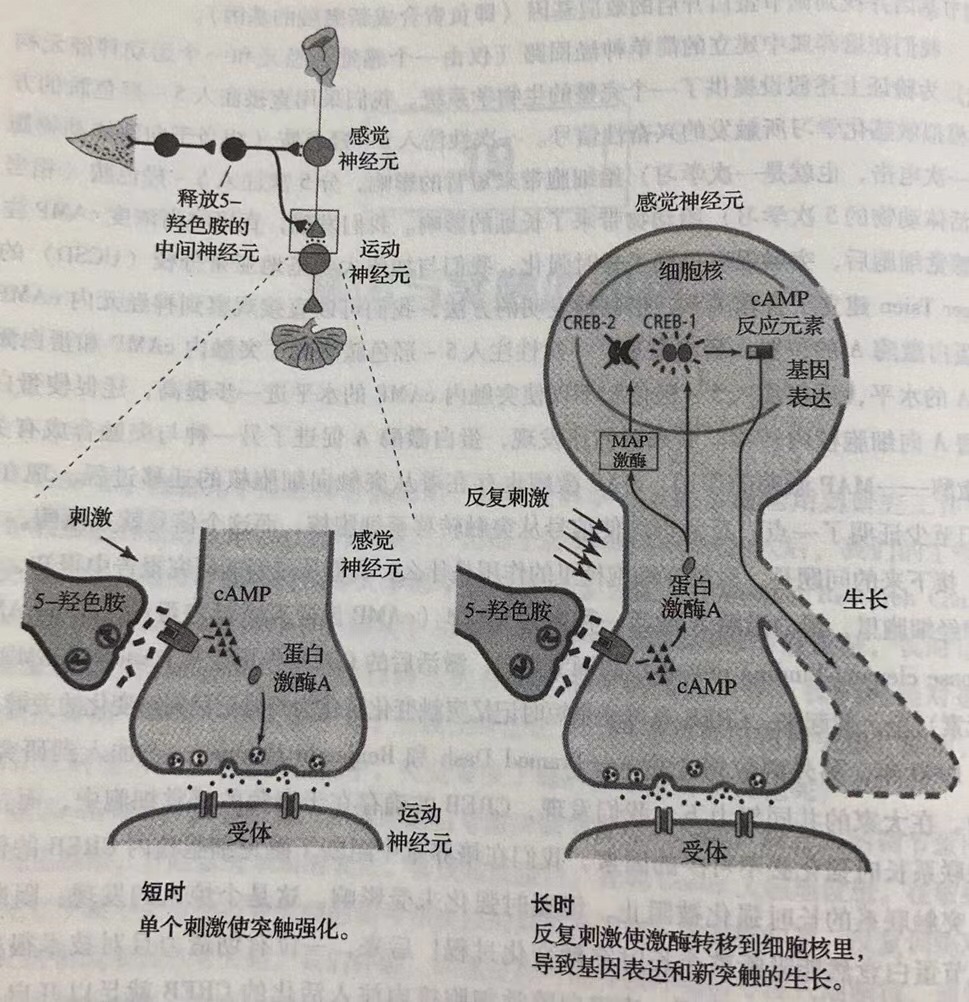
我们的长时记忆研究得到了三条基本原则, 首先,长时记忆的形成需要开启某些基因。其次,不是所有的经验验都能进入长时记忆。为开启长时记忆所需的基因,必须激活CREB-1蛋白,而起抑制作用的CREB-2蛋白必须失活。人们记不住所有的事情,也没人愿意记住所有的事情,所以编码抑制蛋白的基因肯定为经验设立了一个阈限,只有超过这个阈限的经验才能进入到长时记忆中。这就是为什么我们只能记住重要事件的原因,生活中那些普普通通的事情往往都不会记得太久。排除这道阈限使短时记忆的内容顺利进入长时记忆。CREB-1激活的基因编码合成新突触所需的蛋白质。长时记忆需要开启某些基因的事实说明,基因不只是行为的决定者,它们还对环境(如学习)做出反应。
最后,形成长时记忆必须长出新的突触。如果看完这本书后你记住了某些内容,那么你的大脑里一定又多了些突触。这种能力在进化过程中是相对保守的。比如,无论是人类还是较简单的动物,大脑的体表皮层图都受到感觉信息的调节而不断地发生变化。
第五卷
因此,人们对情绪产生达成了一致意见:首先是对某个刺激的无意识的、内隐的评估,随后引起了生理反应,最后出现了或持久或短暂的有意识体验。
杏仁核将情绪的意识体验和情绪的生理反应连接起来,特别是恐惧情绪。
精神分裂症主要有三种临床症状:阳性症状、阴性症状和认知症状。阳性症状,临床中认为最少持续6个月,表现为怪异的行为和心理功能的紊乱,尤其是出现妄想和幻觉状态。这些都是患病期间最显著的症状,患者在此期间不能正确地认知现实世界,也就是说,患者不能真实面对他们的信念和知觉,无法将它们与自己身边世界实际发生的事情作对比。这种对现实世界感知能力欠缺的主要特点为妄想(一种明显违背事实的异常信念,并且此信念不会被能够明显证明其不合理的证据所改变)、幻觉(一种对没有实际发生的外部刺激产生的感知觉,比如听到别人对自己行为的评论)以及不合逻辑的思维(各种想法之间缺乏正常的联系,联系松散或者脱轨,严重时导致思维混乱语无伦次)。
精神分裂症的阴性症状表现为缺少正常的社交人际活动,伴随社交障碍、少言寡语以及情感淡漠等症状。认知症状包括注意力不集中和短时记忆/工作记忆的缺失。认知症状是慢性的、长期的甚至存在于症状缓解期,也是精神分裂症中最难以治疗的方面。
在患病期间,患者主要表现出阴性症状和认知症状:行为异常、与社会隔离、情感淡漠、难以融入社会、少言寡语、注意广度降低以及动机缺乏。
和认知症状都有关联。Carlsson认为,海马和杏仁核与中脑的多巴胺通路中的过多的多巴胺可能导致了阳性症状;而中脑和皮层通路中的过量的多巴胺则可能导致阴性症状和认知症状的产生。研究者们很快发现,缓解精神分裂症状的药物主要针对一种特殊类型的多巴胺受体–D2受体。约翰霍普金斯大学的Solomon Snyder以及多伦多大学的Philip Seeman都发现了抗精神疾病药物的有效性和它们阻滞D2受体的能力之间的显著相关。同时人们也发现,这些药物仅仅对精神分裂的阳性症状有效,减轻了甚至消除了妄想、幻觉以及一些与阴性症状和认知症状联系不大的思维混乱表现。但是很难解释这些药物对于阴性症状和认知症状并无显著效果。
问题之一就是自由意志的本质。除了弗洛伊德的精神决定论(我们认知和情感生活都是无意识的)之外,个人选择,行动自由又是怎么回事呢?
针对这个问题,最为关键的一系列研究是1983年加州大学洛杉矶分校的Benjamin Libet进行的。Libet以德国神经科学家HansKornhuber的发现为起点。在Komhuber的研究中,要求被试移动右手食指,然后用标准尺测量被试移动的长度,同时用头皮电极记录被试的脑电活动。几百次之后,Kornhuber发现,大脑记录的脑电活动都比每一次手指移动早一点点。这就是自由意志!他称这个电位为“准备电位”(readiness po tential),并且发现它比随意移动早1秒发生。
Libet 在Kornhuber的研究基础上,进行了另一个实验,实验中让被试在感到想举起手指时举起手指,他同样用电极记录大脑活动,确认准备电位先于他或她举手指1秒出现。然后发现被试产生想法的时间比准备电位滞后了200毫秒。因此,仅仅通过观察脑电活动,Libet能预期在一个人真正感觉到已经作出决定之前他要做什么。
这个发现引起心理哲学家的疑问:如果选择在我们没有决定之前就已经决定,自由意志在哪儿?难道决定我们活动的感觉只是幻想,只是在发生以后的一种合理化解释?或者决定是自由的,但不是有意识的?如果这样,行为决策就像在知觉中那样,可以反映无意识推理的重要性。Libet提出:开始一个主动的活动加工发生在大脑的无意识部分。但正是在行为开始之前,意识参与进来批准或否决这个行动,这200毫秒就是意识决定是否活动的时间。
无论决定和知觉之间的延迟原因是什么,Libet的发现都提出了一个伦理问题。人们如何对那些没有意识参与的决定负责呢?心理学家Richard Gregory和Vilayanur Ram- achandran对这种争论提出了严格的限制,他们指出:我们的意识可能没有自由意志,但是它有否决的自由。MichaelGazzaniga,认知神经科学的奠基者,也是美国生物伦理学委员会(AmericanCouncil of Bioethics)的成员,也说:“大脑是自动化的,但人类是自由的。”因此,不能仅凭大脑中几个神经回路的活动推断整个神经活动。
第六卷